At the end of 2013, Prof. Stuart Wilson asked me to adapt and put in place for his laboratory a procedure called individual nucleotide resolution UV Cross-Linking and ImmunoPrecipitation (iCLIP) to establish a transcriptome-wide map of the interactions occurring between the major mRNA export factors we had studied so far (i.e. Alyref, Chtop, Nxf1) and their RNA targets in vivo. This technique allows for a transcriptome-wide determination of the RNA-binding sites of a given RNA-Binding Protein (RBP). As its main outputs, it can reveal which RNAs this RBP binds to, how the binding events of that RBP are distributed along its target RNAs, and whether this RBP binds with any sequence specificity. While starting this study with simple questions (« How are the RNA export factors distributed along the RNA, and which RNAs do they bind to? »), I actually made a number of discoveries, not only important for the field of RNA export, but also for the understanding of gene expression in general.
In 2013, the canonical RNA export factors were thought to be mainly deposited at the 5’ end of the RNA, on the Cap-Binding Complex (CBC) as part of the THO-TREX complex 20. Yet, some physical interactions with the Exon Junction Complex (EJC) or functional connections with 3’-end processing had also been reported, which left the topology of RNA export factors on the RNA unclear.
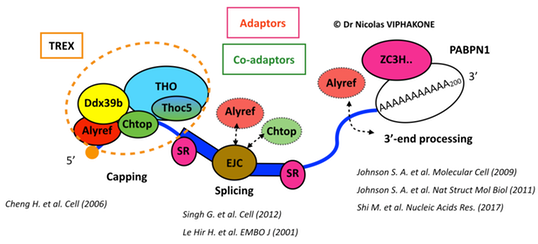
Topology of mRNA export factors on mRNAs, as known in 2013. The CBC is depicted as an orange disc. Dashed shapes and arrows reflect incertainties and traces of interactions known at that time. Some publications reporting the connections between each RNA processing step and RNA export factors are mentioned.
The analysis of our iCLIP data by Dr Ian Sudbury confirmed that Alyref, Chtop and Nxf1 bind to the majority of mRNAs in the cell, which means that they constitute the main pathway for mRNA export. We have also confirmed the enrichment of RNA export factors at the 5’ end of mRNAs. Interestingly, our analysis also revealed that these factors have additional binding patterns along the mRNA and that they differ from one another in that respect. Alyref binds to the 5’ of the RNA and along the CDS while Nxf1 and Chtop also bind along the CDS but predominantly to the 3’UTR.
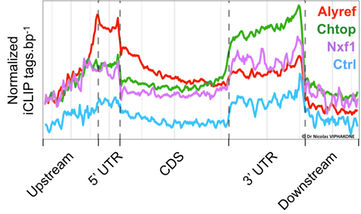
Global distribution of RNA export factors along mRNAs. Ctrl refers to results obtained with negative IP iCLIP experiments.
While we found that RNA export factors globally prefer to bind to spliced RNAs, we have also detected their binding to poorly spliced introns and we have shown that the binding of Alyref regulates at least 20% of those inefficiently spliced introns. Moreover, we found that they are recruited to RNAs that have not yet been subject to 3’-end processing. These results and some others we obtained show that, in humans, RNA export factors are recruited co-transcriptionally, mainly in connection to the splicing process. This is consistent with the fact that I detected RNA export factors mostly attached to the chromatin (see western blot analysis below). Interestingly, these findings suggests that the nucleoplasmic residency time of RNAs covered by RNA export factors is probably short and that the binding of RNA export factors contributes to a strong commitment to nuclear export. However, their presence is clearly not sufficient to trigger export, as we also found them on lncRNAs, even nuclear-retained ones such as XIST. Interestingly, cytoplasmic lncRNAs had comparatively less Nxf1 on them. For further reading, I discussed this at length in the discussion section of the article reporting this study.
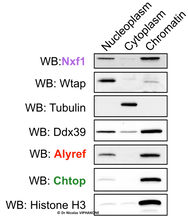
Western blot analysis of a cellular fractionation experiment. The canonical RNA export factors Alyref, Chtop, and Nxf1 are mostly found on the chromatin. Tubulin is a cytoplasmic marker. Histone 3 is a chromatin marker. Wtap is the anchor of the m6A methylase complex and is reproducibly nucleoplasmic is these experiments which were done in the presence of RNase inhibitors.
The presence of Chtop on the chromatin, its strong binding to uncleaved 3’UTRs and its ability to bind to the NTF2L domain of Nxf1 (known interaction site for 3’-end processing regulation factors) led me to test whether Chtop had an influence on poly(A) site choices in vivo. The depletion of Chtop globally affected poly(A) sites usage, mainly triggering a switch from distal to proximal poly(A) sites. Conversely, over-expressing Chtop induced a transcription termination defect at least at the end the CHTOP gene itself. These results and others showed that the RNA export factor Chtop contributes to the choice of cellular poly(A) sites and therefore the length of mRNA 3’UTRs.
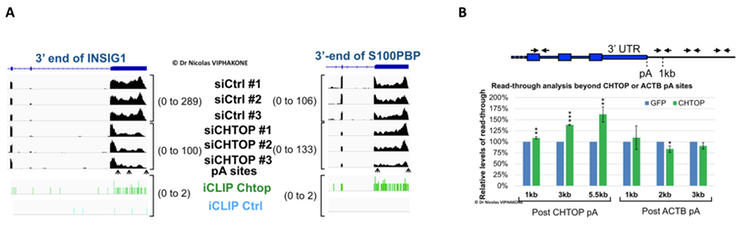
Depletion or over-expression of Chtop affect poly(A) sites choices and transcription termination in vivo. A. Results of an RNA-seq experiment in a context where Chtop has been depleted by RNA interference. B. Over-expression of Chtop from a single copy integrated gene triggers a specific transcriptional read-through beyond the end of the CHTOP gene. Expression of GFP is used as a negative control.
All along the body of the RNA, we found that Alyref sits next to the Exon Junction Complex. This relationship is functionally important because I have also shown that eIF4A3, which anchors the EJC on the RNA, is required for Alyref binding to RNA in vivo and for the nuclear export of a number of RNAs. These results assigned a global function for the biochemical connections described previously between the EJC and RNA export factors 38. While it was known that the EJC binds spliced mRNAs, I also found EJC components on intronless RNAs and poorly spliced introns and showed that the EJC was also required for Alyref to bind to these RNAs. This is consistent with a recent study which found other core EJC components on intronless RNAs in Drosophila 39.
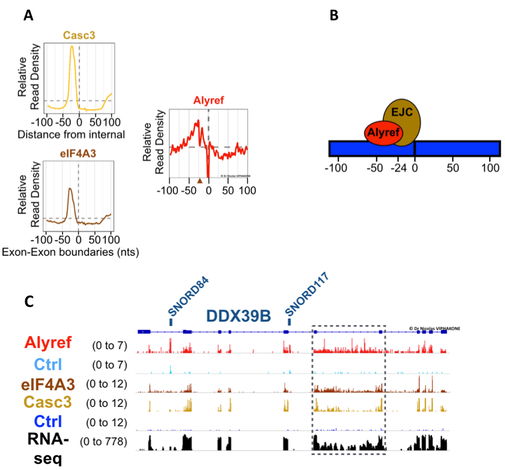
Within the body of mRNAs, Alyref sits next to the Exon Junction Complex. A. Global iCLIP binding distributions of Alyref and EJC components (i.e. Casc3 and eIF4A3) in the vicinity of exon-exon junctions. B. Model describing data seen in A. C. Alyref and EJC subunits RNA binding events along the RNA DDX39B. The dotted box highlights binding of these RBPs to a poorly spliced intron.
Finally, with a series of biochemical experiments, I was able to shed some light on the mechanisms leading to the recruitment of the main RNA export adaptor Alyref on RNAs in the cell. Throughout transcription, the Cap-Binding Complex (CBC) seems to be an initial landing pad for Alyref which is then transferred co-transcriptionally next to the EJC all along the body of the RNA in a manner stimulated by the splicing process (i.e. the reaction itself but also its actors).
Overall, this study allowed us to refine the topology of RNA export factors on mRNAs. It also merged ideas previously thought to oppose each other (i.e. CBC-centric 20 and EJC-centric 38 models) into one unifying model which describes how the canonical RNA export factors are first recruited via the 5’ cap and then are distributed along the RNA. Importantly, my study also revealed how the co-transcriptional recruitment of RNA export factors allows them to influence RNA processing (i.e. here splicing and 3’-end processing) and therefore the final state of the human transcriptome before promoting RNA export.
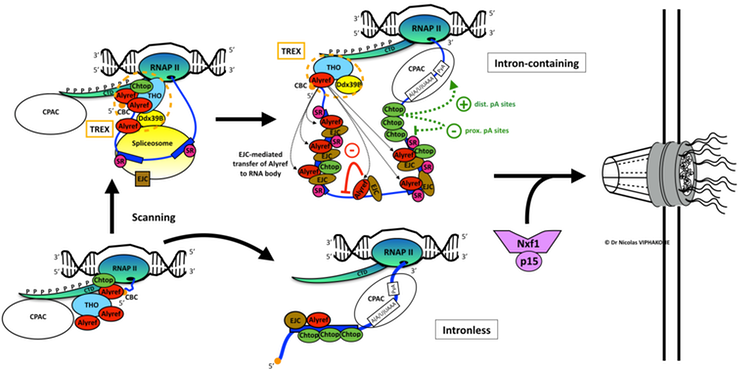
Model of co-transcriptional recruitment of mRNA export factors and its effect on gene expression. Alyref is initially recruited as part of the THO-TREX complex to the Cap-Binding-Complex (CBC, orange disc) at the 5’ end of the RNA. Initial scanning of the RNA by spliceosome/EJC components (and notably eIF4A3) might allow the deposition of RNA export factors onto intronless RNAs but we have shown that this may occur at a lower efficiency than on intron-containing RNAs. On multi-exon transcripts, splicing deposits the EJC. Alyref then transfers from the CBC to exon-exon junctions along the RNA, upstream of the EJC. Intron retention can also trigger atypical binding of the EJC and Alyref, which in turn regulates this phenomenon. As the last exon is produced by the RNA Polymerase II, the presence of Chtop increases and it participates in poly(A) site choice. Ultimately, Alyref and Chtop allow Nxf1 to join the RNP and stimulate nuclear export, as I have shown previously 24,33.
References used:
Preceding references: 1-5, 6-16, 17-27, 28-37.
38. Le Hir, H., Gatfield, D., Izaurralde, E. & Moore, M. J. The exon-exon junction complex provides a binding platform for factors involved in mRNA export and nonsense-mediated mRNA decay. EMBO J 20, 4987–4997 (2001).
39. Choudhury, S. R. et al. Exon junction complex proteins bind nascent transcripts independently of pre-mRNA splicing in Drosophila melanogaster. Elife 5, 720 (2016).